|
||||
|
4. Модели рабочей памяти Из предыдущей главы мы усвоили, что способность сохранять информацию играет базовую роль в решении широкого диапазона интеллектуальных задач. Рабочая память используется для того, чтобы контролировать внимание, запоминать и извлекать из памяти команды, выбирать — какие задачи следует решить сейчас, а какие, более сложные, отложить на потом. Рабочая память имеет ограниченный объем, именно поэтому наша способность обрабатывать информацию и решать задачи имеет ограниченный диапазон. Одна из самых серьезных проблем, с которой мы сталкиваемся в наш век информационного прорыва, — ограниченный объем нашей рабочей памяти. Так что прежде всего нам нужно понять, каким образом сохраняется информация и удастся ли нам преодолеть эти ограничения. Исследовательница Патриция Голдман-Ракич (Йельский университет, США) на протяжении многих лет изучала процессы мозговой деятельности и механизмы рабочей памяти и в числе прочего разрабатывала методику точечных тестов. Она и ее коллеги регистрировали активность нейронов в разных областях мозга у обезьян, чтобы выяснить, как действует механизм рабочей памяти[39]. Выяснилось, что нейроны выключаются, как только инициирован ответ. Это был весьма трудоемкий процесс. К оборудованию подключили усилители и микрофоны. И тогда ученые услышали сигналы электрической активности нейронов — пощелкивание и потрескивание. Эту звуковую карту следовало расшифровать, и задача оказалась не из легких. Однако Патриции Голдман-Ракич удалось охарактеризовать некоторые паттерны. Примечательно, что активность некоторых нейронов проявлялась именно в тот момент, когда информация сохранялась в рабочей памяти. Нейроны начинали активизироваться тогда, когда обезьяна устремляла свой взгляд на точку, которую ей следовало запомнить. Нервные импульсы поступали от нейронов даже после того, как точка исчезала, и до того момента, пока обезьяна переводила свой взгляд на то место, которое запомнила. Подобная активность была названа активностью специфического периода ожидания. Если процесс непрерывной активности прерывался, то обезьяна не могла сохранить информацию. Основная часть нейронов с подобного типа активностью находится в лобной доле, меньшая часть — в теменной. Согласно теории, выдвинутой Патрицией Голдман-Ракич и ее коллегами, информация сохраняется в рабочей памяти благодаря особым нейронам, находящимся в состоянии непрерывной активности. Этот процесс отличается от процесса кодирования информации в долговременной памяти. Связи между нейронами перманентно усиливаются, это требует времени и синтеза новой порции белков. Кодирование информации в рабочей памяти — куда более динамичный процесс. Это оперативный способ сохранения информации, поскольку электрическая активность нервной системы длится всего несколько миллисекунд. Однако это — самый уязвимый способ, поскольку процесс запоминания прекращается, когда обрывается сеть. Теперь вернемся к вопросу об определении разных типов памяти. Чтобы не путаться с терминологией и охарактеризовать специфику функций, которые определяют, что происходит в мозге, нам следует определить рабочую память как «способность сохранять информацию в активном состоянии в течение короткого периода, базирующуюся на непрерывной активности нейронов в лобной доле». Возьмем хотя бы предыдущий пример с автостоянкой, когда мы паркуем машину, чтобы купить пакет молока. Информацию о том, где мы запарковали машину, мы сохраняем в долговременной памяти. Нейроны в лобной доле не участвуют в кодировании информации о местоположении автомобиля, так что пока мы ходим по магазину, эта информация нам не понадобится. 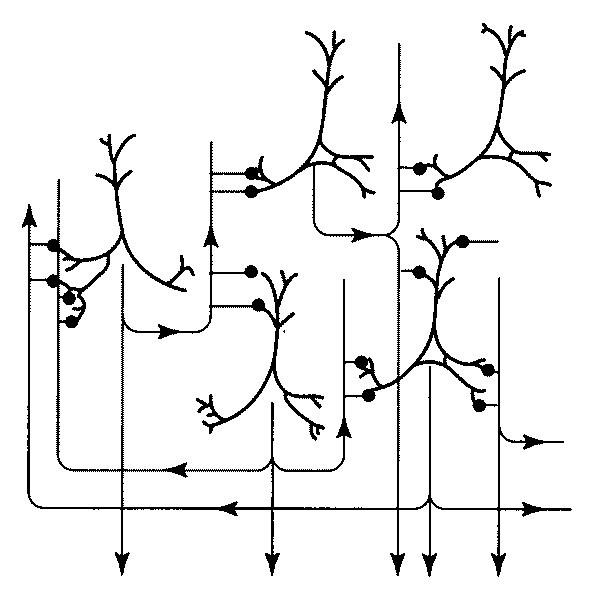 Благодаря компьютерному моделированию можно наблюдать, как происходит процесс активации и как информация сохраняется в памяти путем взаимной активации нейронов Но когда мы ищем молоко в магазине, мы обращаемся за подсказкой к рабочей памяти. Эта информация находится в режиме онлайн, иными словами, она постоянно хранится в нашем сознании. Мы не знаем досконально, каким образом нейронам удается сохранять свою активность на протяжении всего периода. На этот счет существует гипотеза — есть рекуррентные петли, то есть сети из нейронов, которые сохраняют состояние активности путем обмена импульсами. В последние годы ученые, исследующие эти механизмы, добились успехов благодаря методу компьютерного моделирования. Компьютерные модели воспроизводят процесс активизации отдельных нейронов[40]. Виртуальные нейроны связываются друг с другом, образуя сеть. Затем исследователи изучают, при каких условиях возникает и поддерживается состояние активности. Выясняется, что необходим баланс между процессами стимуляции и торможения. Чрезмерное торможение приводит к тому, что нейронная активность обнуляется, соответственно, теряется и сохраненная информация. И напротив, чрезмерная активность приводит к виртуальной эпилепсии мозга. Информация в теменной доле В 1990-е годы знания о функционировании рабочей памяти начали активно пополняться, когда благодаря методу позитронно-эмиссионной томографии (ПЭТ) ученые получили возможность измерять уровень мозгового кровотока, в то время как испытуемые решали задачи, за которые отвечает рабочая память. Наблюдая за процессом активизации лобной доли, ученые пришли к тем же выводам, что и исследователи функции лобной доли у обезьян и травм лобной доли у людей. Позитронно-эмиссионный томограф дает более детальную информацию, и исследователи получили возможность выделить те области, которые сохраняют визуальную информацию в рабочей памяти, и те области, которые активизируются в процессе запоминания вербальной информации. Позитронно-эмиссионный томограф позволял фиксировать информацию каждую минуту[41]. В середине 1990-х годов ученые начали использовать функциональный магнитно-резонансный сканер, чтобы получать снимки мозговой активности примерно каждую секунду. Техника с более высоким разрешением позволяет различать активность на разных стадиях — на самой ранней, когда происходит ознакомление с объектом, на стадии сохранения информации в рабочей памяти и на заключительной стадии, когда мы реагируем на ту или иную информацию[42]. Многие исследования ставили перед собой задачу — проанализировать степень активности мозга, характерную для стадии сохранения информации в рабочей памяти. Ученые убедились в том, что информация сохраняется за счет непрерывной активности нейронов в лобной доле. Эти опыты дали очень серьезные результаты. И кстати, выяснилось, что не только лобная доля сохраняет непрерывную активность на стадии запоминания, но также и теменная доля. Соотношение памяти и внимания Сравнивая данные тестов на произвольное внимание с данными тестов на рабочую память, мы убедились в том, что рабочая память и управление вниманием связаны, и это согласуется с некоторыми психологическими концепциями. Но активируется ли при этом одна и та же система мозга? Вот в чем вопрос. В одном из исследований мозговой деятельности в процессе активации рабочей памяти Клейтон Кертис и Марк Д'Эспозито (университет Беркли, штат Калифорния, США) использовали те же самые точечные тесты, которые ранее использовались с обезьянами[43]. Ученые работали с группой из пятнадцати человек. Мозговая активность каждого из них измерялась в течение 45 минут, а снимки мозговой активности делались каждую секунду. Эти эксперименты потребовали воли и выносливости не только от испытуемых, которые помещались в камеру томографа и запоминали позиции из точек на протяжении 45 минут, но и от ученых, которым предстояло обработать результаты более чем 40 тысяч снимков мозга. После статистического анализа этих изображений Кертис и Д'Эспозито зафиксировали активность в теменной доле (sulcus intraparietalis), в верхней лобной извилине (gyrus frontalis superior), а также в передней части лобной доли (gyrus frontalis media). Примечательно, что в экспериментах Познера на произвольное внимание также активизировались первые две области. Результаты исследований мозга подтверждают гипотезу о том, что рабочая память связана с управлением вниманием. Это может означать, что процессы запоминания точки и запоминания, куда следует направить свое внимание в ожидании появления точки, ничем не отличаются. И все же следует подчеркнуть, что процессы активизации рабочей памяти и произвольного внимания совпадают не полностью. Во многих случаях, когда рабочая память выполняет определенные задания, в лобной доле наблюдается активация, которая не всегда проявляется при решении задач на произвольное внимание. Какую роль играет в данном случае процесс активации — до конца не ясно. Но так или иначе на нашей карте мозговой активности по-прежнему много белых, или, по крайней мере, серых пятен. Мы по-прежнему не знаем, как функционируют передние части лобной доли. Возможно, в процессе активации передней части лобной доли взаимодействие между верхними частями лобной доли и теменной доли стабилизируется. 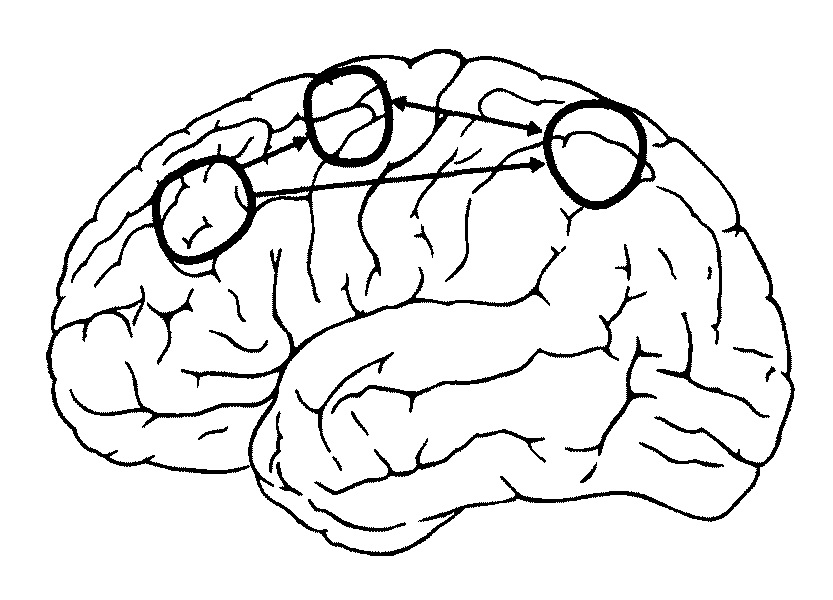 Кругами обведены области мозга, которые активизируются, когда информация сохраняется в рабочей памяти. Область в теменной доле и верхняя область лобной доли находятся в состоянии непрерывной активности в процессе сохранения пространственной информации. Эти области идентичны тем областям, которые активизируются при произвольном внимании. Область в лобной доле активизируется, когда включается рабочая память, но не всегда активизируется при контроле внимания. Стрелки показывают, как области сообщаются друг с другом в процессе функционирования рабочей памяти[44]. Как кодируется информация И все-таки нам крайне важно выяснить — каким образом нейронам удается сохранять активность на стадии запоминания без внешних стимулов? Возможно, этот механизм запускают внутренние связи между нейронами. Другой важный вопрос — какого типа информация кодируется в процессе непрерывной активности? Какая роль отведена этому процессу? Подобные вопросы возникали и раньше, когда проводились исследования по изучению долговременной памяти. Согласно одной из теорий, определенные нейроны отвечают за специфическую память. Речь идет о так называемой «теории бабушкиных клеток». По этой теории, у нас есть специфическая клетка, которая активируется каждый раз, когда мы видим свою бабушку. Что касается рабочей памяти, то на этот счет существует теория сенсорной информации, которая передается от тыльных областей мозга к специализированным нейронам лобной доли, и эта теория в чем-то сходна с «теорией бабушкиных клеток». Непрерывная активность специфических нейронов лобной доли, таким образом, позволяет обезьяне запомнить, что она видела точку, расположенную на 90 градусов справа. Активизируясь, близлежащая клетка передает память о точке на 120 градусов справа и так далее. Согласно другой теории, информация о разных стимулах может кодироваться на той же частоте, на которой активизируются нейроны. Но есть и другие теории, согласно которым информация не всегда считывается благодаря активности нейронов в лобной доле. Определенные нейроны демонстрируют активность рабочей памяти независимо от того, какие стимулы запоминаются. Поскольку они кодируют разные модели чувств, такие как звуковая и визуальная информация, их можно назвать мультимодальными — это своего рода универсальные нейроны[45]. Все эти вопросы могли бы показаться чрезмерно педантичными и академическими, и, возможно, даже не имеющими отношения к теме, если бы не исключительная заинтересованность (в том числе и моя личная) в классификации различных видов нейронов в лобных долях. Но способ кодирования информации, бесспорно, влияет и на способ организации информационных потоков в мозге. Если разные клетки в лобной доле кодируют разные стимулы, то это свидетельствует о параллельной организации информационного потока. 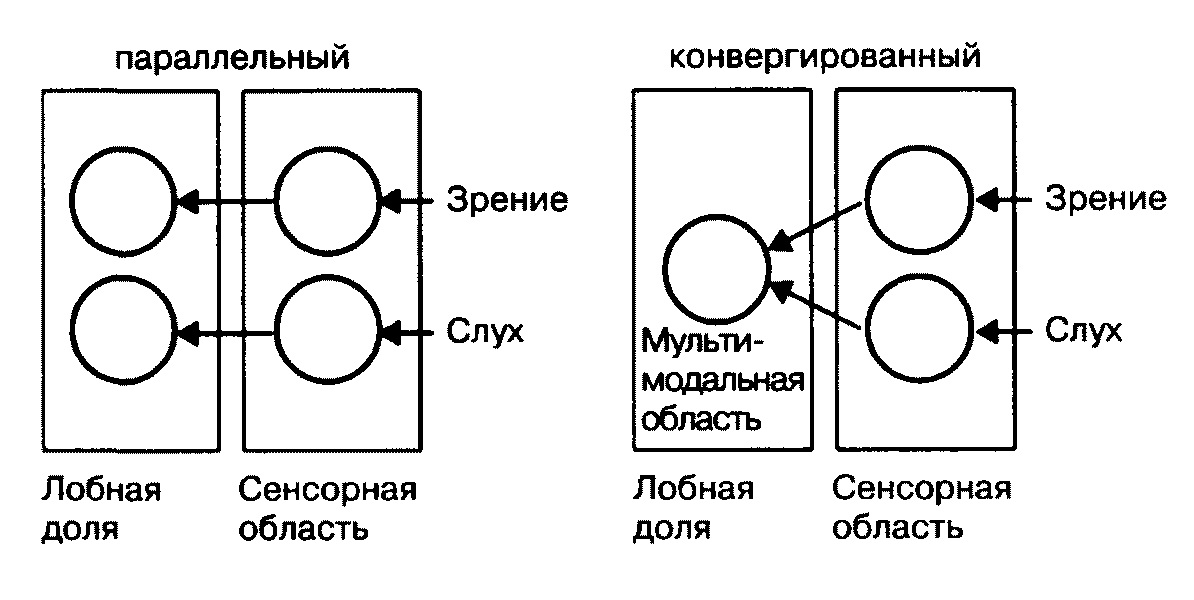 Схема параллельного и конвергированного потоков информации при активации рабочей памяти Патриция Голдман-Ракич, которая отстаивала эту модель, утверждала, что рабочая память опирается на параллельные системы, каждая из которых обрабатывает свой тип информации. С другой стороны, если существуют мультимодальные клетки, вовлеченные в рабочую память, то они должны получать информацию от разных сенсорных нейронов в тыловой части мозга, и это уже конвергированный поток информации[46]. Некоторые исследования рабочей памяти все же дают повод для дискуссий о том, как кодируется информация. В одном из экспериментов измерялся уровень мозговой активности, например в процессе запоминания тонов и уровня света — испытуемым предлагалось запомнить и сравнить, насколько ярок свет от лампы сейчас и потом, спустя несколько секунд. Некоторые области мозга активизировались независимо от того, какой тип информации человек запоминал, иными словами, выполняли роль мультимодальных областей рабочей памяти. А это обстоятельство уже противоречит теории Патриции Голдман-Ракич о параллельных системах восприятия разного типа информации[47]. Многие дальнейшие исследования также подтвердили эти выводы. Как интерпретировать эти данные? Сам факт, что мы обнаружили определенные области, где конвергируется обработка информации, имеет важные функциональные последствия. Параллельная организация функций кажется более гибкой и менее уязвимой для сбоев, к тому же она позволяет вмещать большее количество информации. Вспомним многопроцессорные компьютеры, которые превосходят однопроцессорные. А любые стыки и соединения уязвимы и подвержены эффекту «узкого бутылочного горла». Можно сделать вывод: когда на мозг каменного века обрушивается поток информации, то ограниченный объем рабочей памяти становится серьезной проблемой. Удастся ли нам выяснить, какие факторы ограничивают нашу рабочую память, какую роль в этом процессе играют мультимодальные области мозга? Или, возможно, существует отдельная область мозга, которая ограничивает объем рабочей памяти или нашу способность решать те или иные задачи? Примечания:3 Hallowell, E. Overloaded Circuits: Why Smart People Underperform. Harvard Business Review. 2005. Jan. 01. 4 Miller, G.A. The magical number seven, plus-or-minus two or some limits on our capacity for processing information. Psychological Review. 1956. 63:81–97. 39 Одна из самых цитируемых работ на тему активности нейронов и задачах рабочей памяти: Funahashi, S., Bruce, C.J. & Goldman-Rakic, P.S. Mnemonic coding of visual space in the monkey's dorsolateral prefrontal cortex. Journal of Neurophysiology. 1989, 61:331–349. См. также более ранние работы на эту же тему: Fuster, J.M. & Alexander, G.E. Neuron activity related to short-term memory. Science, 1971. 173:652–654. 40 О компьютерных моделях рабочей памяти см: Wang, X.-J. Synaptic reverberation underlying mnemonic persistent activity.Trends in Neuroscience. 2001.24; Tegner, J., Compte, A. & Wang, X.J. The dynamical stability of reverberatory neural circuits. Biological Cybernetics. 2002. 87:471–481. 41 Результаты более ранних исследований рабочей памяти на позитронно-эмиссионном томографе обобщены в работах: Paulesu, E., Frith, C.D. & Frackowiak, R.S.J. The neural correlates of the verbal component of working memory. Nature. 1993. 362:342–345; Jonides, J., Smith, E.E., Koeppe, R.A., Awh, E., Minoshima, S. & Mintun, M.A. Spatial working memory in humans as revealed by PET. Nature. 1993. 363: 623–625. 42 О непрерывной активности нейронов в процессе использования функциональной магнитно-резонансной томографии см.: Cohen, J.D., Pearstein, W.M., Braver, T.S., Nystrom, L.E., Noll, D.C., Jonides, J. & Smith, E.E.Temporal dynamics of brain activation during a working memory task. Nature. 1997.386; Courtney, SM., Ungerleider, L.G., Keil, K. & Haxby, J.V. Transient and sustained activity in a distributed neural system for human working memory. Nature. 1997. 386:608–611. 43 О постоянной активности в процессе проведения точечных тестов см.: Curtis, C.E., Rao.V.Y. & D'Esposito, M. Maintenance of spatial and motor codes during oculomotor delayed response tasks. Journal of Neuroscience.2004. 24:3944–3952. 44 Иллюстрация заимствована из источника: Curtis, C.E. & D'Esposito, M. Persistent activity in the prefrontal cortex during working memory. Trends in Cognitive Sciences. 2003. 7:415–423. 45 Об исследованиях, которые поддерживают теорию специализированных нейронов см.: Funahashi, S., Bruce, C.J. & Goldman-Rakic, P.S. Mnemonic coding of visual space in the monkey's dorsolateral prefrontal cortex. Journal of Neurophysiology. 1989. 61. 46 Исследования о мультимодальных клетках: Quintana, J. & Fuster, J.M. Mnemonic and predictive functions of cortical neurons in a memory task. Neuroreport, 1992. 3:721–724. Для сравнения см.: Fuster, J.M. Memory in the cerebral cortex. Cambridge, Massachusetts: 1995. MIT Press. 47 Теория о параллельных системах внимания изложена в следующих работах: Goldman-Rakic, P.S. Topography of cognition: parallel distributed networks in primate association cortex. Annual Reviews of Neuroscience. 1988. 11:137–156; Duncan, J. & Owen, A.M. Common regions of the human frontal lobe recruited by diverse cognitive demands. Trends in Neurosciences. 2000. 23:475–483; Hautzel, H., Mottaghy, F.M., Schmidt, D., Zemb, M., Shah, N.J., Muller-Gartner, H.W. & Krause, B.J. Topographic segregation and convergence of verbal, object, shape and spatial working memory in humans. Neuroscience Letters. 2002. 323:156–160; Curtis, C.E. & D'Esposito, M.Persistent activity in the prefrontal cortex during working memory. Trends in Cognitive Sciences. 2003. 7:415–423. |
|
|||
|
Главная | В избранное | Наш E-MAIL | Добавить материал | Нашёл ошибку | Наверх |
||||
|
|
||||